NEURON
| LE NEURONE - UNITÉ FONCTIONNELLE DU SYSTÈME NERVEUX
| L'intégrité d'un organisme vivant nécessite l'activité coordonnée des cellules qui le composent. Cette coordination est assurée par des mécanismes de communication intercellulaire impliquant soit des cellules d'un même tissu soit des cellules pouvant être très éloignées, appartenant à des tissus différents. La communication intercellulaire est assurée par deux grands systèmes : le système hormonal (communication humorale) et le système nerveux, qui transmet l'information au moyen de prolongements cellulaires. Ces deux systèmes sont étroitement liés (contrôle hypothalamique des sécrétions hormonales) et la distinction entre les deux systèmes peut être arbitraire (neurones sécrétoires de l'hypothalamus : ADH, ocytocine). Le neurone, unité fonctionnelle du système nerveux, est responsable de l'émission et de la propagation du message nerveux.
Les neurones ont des formes très diverses et leur morphologie (dimension et orientation des champs dendritiques, taille du soma ) joue un rôle fondamental dans leurs capacités d'intégration et de traitement de l'information. Les neurones sont amitotiques (incapables de se reproduire) et ont donc une longévité extrême. De plus, la vitesse de leur métabolisme est exceptionnellement élevée. Ils requièrent de ce fait un approvisionnement constant et abondant en oxygène et en glucose (mort cérébrale en quelques minutes en cas d'anoxie cérébrale).
| LES NEURONES ONT UN CORPS CELLULAIRE D' OU PARTENT DEUX TYPES DE PROLONGEMENTS : LES DENDRITES ET L'AXONE
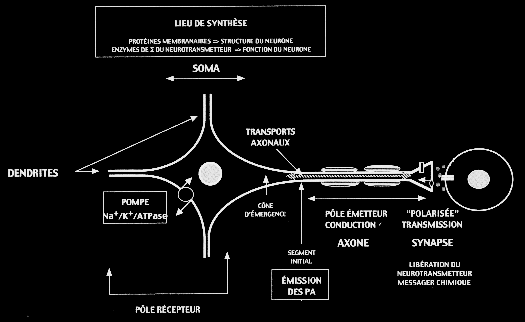
| L'ARBRE SOMATO-DENDRITIQUE REPRÉSENTE LE PÔLE RÉCEPTEUR DU NEURONELe corps cellulaire ou soma contient le noyau cellulaire et le cytoplasme qui l'entoure ou périkaryon. Il est de forme très variable (cellules pyramidales du cortex, somas ovoïdes des cellules de Purkinje, motoneurones multipolaires de la moelle épinière ). Il assure la synthèse des constituants nécessaires à la structure et aux fonctions du neurone et ce, pendant toute la vie de l'individu. Il présente plusieurs prolongements, qui émergent du corps cellulaire et s'arborisent plus ou moins abondamment. Ces prolongements leur permettent d'établir des contacts (synapses) avec d'autres neurones ou d'autres cellules de l'organisme (récepteurs sensoriels, cellules musculaires). Ces prolongements sont de deux types : les dendrites et l'axone, qui se distinguent sur la base de critères morphologiques, métaboliques et fonctionnels. Les dendrites, lorsqu'elles émergent du soma, sont de simples prolongements du périkaryon (troncs dendritiques). Ces troncs dendritiques (1-9) se divisent successivement pour donner un arbre dendritique, dont les caractéristiques morphologiques sont propres à chaque population neuronale. Les contours irréguliers des dendrites sont dus à la présence, à leur surface, d'épines dendritiques, expansions latérales reliées aux branches dendritiques par un pédicule plus ou moins long. Les neurones sont ainsi dits "épineux" ou "lisses". Les dendrites et le soma reçoivent de très nombreux contacts synaptiques venant d'autres neurones et constituent la principale surface de réception du neurone. Ils intègrent les messages afférents et génèrent, en réponse à ces messages, des signaux électriques (potentiels post-synaptiques). L'AXONE ET SES COLLATÉRALES REPRÉSENTENT LE PÔLE ÉMETTEUR DU NEURONEL'axone se différencie des dendrites par son aspect lisse et son diamètre uniforme le long de son trajet. Il prend généralement son origine au niveau d'une expansion conique du soma : le cône d'émergence. Le segment initial de l'axone est de diamètre plus petit que l'axone lui-même. Il se divise ensuite en une ou plusieurs collatérales d'axone, dont certaines retournent vers le corps cellulaire (collatérales récurrentes). L'axone et ses collatérales peuvent être recouverts d'une gaine de myéline, formée par les cellules gliales. La longueur de l'axone est très variable d'un neurone à l'autre. On distingue ainsi :
Pour certains neurones, l'axone et ses collatérales se terminent par une arborisation terminale, soit par de très nombreuses et très fines branches dont l'extrémité renflée effectue des contacts synaptiques avec les cellules cibles : les boutons terminaux. Pour d'autres, l'axone et ses collatérales présentent le long de leur trajet terminal des renflements ou varicosités qui établissent des contacts synaptiques avec les cellules cibles : les boutons en passant. C'est au niveau du segment initial, que sont générés les potentiels d'action en réponse aux informations synaptiques transmises par l'arbre somato-dendritique. L'axone est ensuite capable de conduire les potentiels d'action sur de longue distance sans perte d'amplitude. Les terminaisons axonales, en réponse à l'arrivée des potentiels d'action, libèrent ou non les neurotransmetteurs qu'elles renferment. Cette libération est localisée au niveau des synapses. L'axone est donc bien le pôle émetteur du neurone. Cependant, certaines régions de l'axone (segment initial, noeud de Ranvier, terminaison axonale) peuvent être également des régions réceptrices de contacts synaptiques venant d'autres neurones (synapses axo-axoniques : cellules de Purkinje).
| LES NEURONES SE CARACTÉRISENT PAR UNE DISTRIBUTION NON HOMOGÈNE DES ORGANITES CELLULAIRES ENTRE LEURS SOMAS ET LEURS PROLONGEMENTS
| LES ÉLÉMENTS DU CYTOSQUELETTE SONT TRÈS ABONDANTS DANS LES NEURONES. ILS JOUENT UN RÔLE DANS LE TRANSPORT DES ORGANITES.Le neurone renferme dans son cytoplasme un réseau très dense de filaments. Le réseau de filaments forme le squelette du neurone : il lui donne une certaine rigidité, notamment au niveau de ses prolongements. Les filaments sont des polymères de protéines qui forment un réseau tridimensionnel, structurant l'espace intracellulaire et formant des architectures spécialisées selon les différentes régions du neurone. On distingue 3 types de filaments : 1. les microtubules - 2. les microfilaments - 3. les neurofilaments. Les microtubules et les microfilaments sont des polymères labiles et dynamiques, qui se polymérisent et se dépolymérisent très rapidement. Les microtubules sont des structures polaires : l'extrémité qui se polymérise est [+], celle qui se dépolymérise est [-]. Ils servent de support aux transports axonaux rapides (antérograde & rétrograde) et les protéines qui leur sont associées jouent le rôle de moteur dans ces transports. LE SOMA EST LE PRINCIPAL LIEU DE SYNTHÈSE DES MACROMOLÉCULES DU NEURONELe soma est le lieu de synthèse des macromolécules du neurone, et ce pendant toute la vie de l'individu. Le nucléole, lieu de synthèse des ribosomes, a une taille particulièrement importante, liée à la forte activité de synthèse protéique des neurones. LES DENDRITES RENFERMENT DES RIBOSOMES LIBRESDans les dendrites, outre les microtubules allongés parallèlement à leurs grands axes, on trouve un réticulum endoplasmique lisse, des mitochondries allongées et des ribosomes libres. Du fait de la présence des ribosomes libres, les dendrites sont capables de synthétiser, du moins en partie, leurs propres protéines (protéines du cytosquelette). L'AXONE EST, EN REVANCHE, INCAPABLE D'ASSURER SA PROPRE SYNTHÈSE PROTÉIQUECelui-ci est dépourvu de ribosomes (libres ou associés au réticulum endoplasmique). Il ne peut donc pas renouveler lui-même ses propres macromolécules constitutives ni assurer la synthèse du ou des neurotransmetteurs qu'il libère. Il renferme de très nombreux éléments du cytosquelette, des mitochondries allongées et des vésicules de transport.
| DES TRANSPORTS AXONAUX PERMETTENT LE TRANSFERT BIDIRECTIONNEL DES MOLÉCULES DU SOMA AUX TERMINAISONS AXONALES
| On distingue deux transports rapides (antérograde & rétrograde), un transport antérograde lent et le transport des mitochondries. LE TRANSPORT ANTÉROGRADE RAPIDEIl consiste en un transport de vésicules (40-60 nm) le long des microtubules de l'axone à une vitesse de 100 à 400 mm par jour. Ces vésicules, formées dans le soma à partir de l'appareil de Golgi, contiennent des protéines nécessaires au renouvellement de la membrane de l'axone, des enzymes de synthèse du ou des neurotransmetteur(s) libéré(s) par la terminaison axonale et le précurseur du neurotransmetteur (peptide). Ce transport est indépendant du type d'axone. LE TRANSPORT ANTÉROGRADE LENTIl consiste essentiellement en un transport du cytosquelette à une vitesse de 0,1 à 2 mm par jour. Il assure le renouvellement de 80% des protéines totales de l'axone. Ce transport est spécifique du type d'axone. Son mécanisme reste inconnu. LE TRANSPORT RÉTROGRADE RAPIDEIl consiste en un transport de corps plurivésiculaires (100-300 nm) le long des microtubules de l'axone à une vitesse de 100 à 400 mm par jour. Ces corps plurivésiculaires permettent le retour des molécules membranaires vers le soma afin qu'elles y soient dégradées (élimination des déchets). De plus, des molécules, absorbées au niveau des terminaisons, peuvent remonter jusqu'au soma et informer la cellule sur ce qu'il se passe à la périphérie ou jouer un rôle trophique (NGF) (communication cellulaire). LE TRANSPORT DES MITOCHONDRIESIl consiste en un transport des mitochondries nouvellement formées dans le soma jusqu'aux terminaisons axonales à une vitesse de 10 à 40 mm par jour. Ce transport consiste en des mouvements saccadés de va-et-vient. Les mitochondries en dégénérescence (corps multilamellaires) remontent vers le corps cellulaire pour y être dégradées. Le mécanisme intime de ce transport reste inconnu.
| 
Iconographie personnelle - Dr. D. Rose
| Le neurone est une cellule excitable et sécrétrice qui présente une extrême régionalisation de ses fonctions :
|
|
L'intégrité d'un organisme vivant nécessite l'activité coordonnée des cellules qui le composent. Cette coordination est assurée par des mécanismes de communication intercellulaire impliquant soit des cellules d'un même tissu soit des cellules pouvant être très éloignées, appartenant à des tissus différents. La communication intercellulaire est assurée par deux grands systèmes : le système hormonal (communication humorale) et le système nerveux, qui transmet l'information au moyen de prolongements cellulaires. Ces deux systèmes sont étroitement liés (contrôle hypothalamique des sécrétions hormonales) et la distinction entre les deux systèmes peut être arbitraire (neurones sécrétoires de l'hypothalamus : ADH, ocytocine). Le neurone, unité fonctionnelle du système nerveux, est responsable de l'émission et de la propagation du message nerveux.
Les neurones ont des formes très diverses et leur morphologie (dimension et orientation des champs dendritiques, taille du soma ) joue un rôle fondamental dans leurs capacités d'intégration et de traitement de l'information. Les neurones sont amitotiques (incapables de se reproduire) et ont donc une longévité extrême. De plus, la vitesse de leur métabolisme est exceptionnellement élevée. Ils requièrent de ce fait un approvisionnement constant et abondant en oxygène et en glucose (mort cérébrale en quelques minutes en cas d'anoxie cérébrale). | ||||||||||
LES NEURONES ONT UN CORPS CELLULAIRE D' OU PARTENT DEUX TYPES DE PROLONGEMENTS : LES DENDRITES ET L'AXONE | ||||||||||
L'ARBRE SOMATO-DENDRITIQUE REPRÉSENTE LE PÔLE RÉCEPTEUR DU NEURONELe corps cellulaire ou soma contient le noyau cellulaire et le cytoplasme qui l'entoure ou périkaryon. Il est de forme très variable (cellules pyramidales du cortex, somas ovoïdes des cellules de Purkinje, motoneurones multipolaires de la moelle épinière ). Il assure la synthèse des constituants nécessaires à la structure et aux fonctions du neurone et ce, pendant toute la vie de l'individu. Il présente plusieurs prolongements, qui émergent du corps cellulaire et s'arborisent plus ou moins abondamment. Ces prolongements leur permettent d'établir des contacts (synapses) avec d'autres neurones ou d'autres cellules de l'organisme (récepteurs sensoriels, cellules musculaires). Ces prolongements sont de deux types : les dendrites et l'axone, qui se distinguent sur la base de critères morphologiques, métaboliques et fonctionnels. Les dendrites, lorsqu'elles émergent du soma, sont de simples prolongements du périkaryon (troncs dendritiques). Ces troncs dendritiques (1-9) se divisent successivement pour donner un arbre dendritique, dont les caractéristiques morphologiques sont propres à chaque population neuronale. Les contours irréguliers des dendrites sont dus à la présence, à leur surface, d'épines dendritiques, expansions latérales reliées aux branches dendritiques par un pédicule plus ou moins long. Les neurones sont ainsi dits "épineux" ou "lisses". Les dendrites et le soma reçoivent de très nombreux contacts synaptiques venant d'autres neurones et constituent la principale surface de réception du neurone. Ils intègrent les messages afférents et génèrent, en réponse à ces messages, des signaux électriques (potentiels post-synaptiques). L'AXONE ET SES COLLATÉRALES REPRÉSENTENT LE PÔLE ÉMETTEUR DU NEURONEL'axone se différencie des dendrites par son aspect lisse et son diamètre uniforme le long de son trajet. Il prend généralement son origine au niveau d'une expansion conique du soma : le cône d'émergence. Le segment initial de l'axone est de diamètre plus petit que l'axone lui-même. Il se divise ensuite en une ou plusieurs collatérales d'axone, dont certaines retournent vers le corps cellulaire (collatérales récurrentes). L'axone et ses collatérales peuvent être recouverts d'une gaine de myéline, formée par les cellules gliales. La longueur de l'axone est très variable d'un neurone à l'autre. On distingue ainsi :
Pour certains neurones, l'axone et ses collatérales se terminent par une arborisation terminale, soit par de très nombreuses et très fines branches dont l'extrémité renflée effectue des contacts synaptiques avec les cellules cibles : les boutons terminaux. Pour d'autres, l'axone et ses collatérales présentent le long de leur trajet terminal des renflements ou varicosités qui établissent des contacts synaptiques avec les cellules cibles : les boutons en passant. C'est au niveau du segment initial, que sont générés les potentiels d'action en réponse aux informations synaptiques transmises par l'arbre somato-dendritique. L'axone est ensuite capable de conduire les potentiels d'action sur de longue distance sans perte d'amplitude. Les terminaisons axonales, en réponse à l'arrivée des potentiels d'action, libèrent ou non les neurotransmetteurs qu'elles renferment. Cette libération est localisée au niveau des synapses. L'axone est donc bien le pôle émetteur du neurone. Cependant, certaines régions de l'axone (segment initial, noeud de Ranvier, terminaison axonale) peuvent être également des régions réceptrices de contacts synaptiques venant d'autres neurones (synapses axo-axoniques : cellules de Purkinje). | ||||||||||
LES NEURONES SE CARACTÉRISENT PAR UNE DISTRIBUTION NON HOMOGÈNE DES ORGANITES CELLULAIRES ENTRE LEURS SOMAS ET LEURS PROLONGEMENTS | ||||||||||
LES ÉLÉMENTS DU CYTOSQUELETTE SONT TRÈS ABONDANTS DANS LES NEURONES. ILS JOUENT UN RÔLE DANS LE TRANSPORT DES ORGANITES.Le neurone renferme dans son cytoplasme un réseau très dense de filaments. Le réseau de filaments forme le squelette du neurone : il lui donne une certaine rigidité, notamment au niveau de ses prolongements. Les filaments sont des polymères de protéines qui forment un réseau tridimensionnel, structurant l'espace intracellulaire et formant des architectures spécialisées selon les différentes régions du neurone. On distingue 3 types de filaments : 1. les microtubules - 2. les microfilaments - 3. les neurofilaments. Les microtubules et les microfilaments sont des polymères labiles et dynamiques, qui se polymérisent et se dépolymérisent très rapidement. Les microtubules sont des structures polaires : l'extrémité qui se polymérise est [+], celle qui se dépolymérise est [-]. Ils servent de support aux transports axonaux rapides (antérograde & rétrograde) et les protéines qui leur sont associées jouent le rôle de moteur dans ces transports. LE SOMA EST LE PRINCIPAL LIEU DE SYNTHÈSE DES MACROMOLÉCULES DU NEURONELe soma est le lieu de synthèse des macromolécules du neurone, et ce pendant toute la vie de l'individu. Le nucléole, lieu de synthèse des ribosomes, a une taille particulièrement importante, liée à la forte activité de synthèse protéique des neurones. LES DENDRITES RENFERMENT DES RIBOSOMES LIBRESDans les dendrites, outre les microtubules allongés parallèlement à leurs grands axes, on trouve un réticulum endoplasmique lisse, des mitochondries allongées et des ribosomes libres. Du fait de la présence des ribosomes libres, les dendrites sont capables de synthétiser, du moins en partie, leurs propres protéines (protéines du cytosquelette). L'AXONE EST, EN REVANCHE, INCAPABLE D'ASSURER SA PROPRE SYNTHÈSE PROTÉIQUECelui-ci est dépourvu de ribosomes (libres ou associés au réticulum endoplasmique). Il ne peut donc pas renouveler lui-même ses propres macromolécules constitutives ni assurer la synthèse du ou des neurotransmetteurs qu'il libère. Il renferme de très nombreux éléments du cytosquelette, des mitochondries allongées et des vésicules de transport. | ||||||||||
DES TRANSPORTS AXONAUX PERMETTENT LE TRANSFERT BIDIRECTIONNEL DES MOLÉCULES DU SOMA AUX TERMINAISONS AXONALES | ||||||||||
|
On distingue deux transports rapides (antérograde & rétrograde), un transport antérograde lent et le transport des mitochondries. LE TRANSPORT ANTÉROGRADE RAPIDEIl consiste en un transport de vésicules (40-60 nm) le long des microtubules de l'axone à une vitesse de 100 à 400 mm par jour. Ces vésicules, formées dans le soma à partir de l'appareil de Golgi, contiennent des protéines nécessaires au renouvellement de la membrane de l'axone, des enzymes de synthèse du ou des neurotransmetteur(s) libéré(s) par la terminaison axonale et le précurseur du neurotransmetteur (peptide). Ce transport est indépendant du type d'axone. LE TRANSPORT ANTÉROGRADE LENTIl consiste essentiellement en un transport du cytosquelette à une vitesse de 0,1 à 2 mm par jour. Il assure le renouvellement de 80% des protéines totales de l'axone. Ce transport est spécifique du type d'axone. Son mécanisme reste inconnu. LE TRANSPORT RÉTROGRADE RAPIDEIl consiste en un transport de corps plurivésiculaires (100-300 nm) le long des microtubules de l'axone à une vitesse de 100 à 400 mm par jour. Ces corps plurivésiculaires permettent le retour des molécules membranaires vers le soma afin qu'elles y soient dégradées (élimination des déchets). De plus, des molécules, absorbées au niveau des terminaisons, peuvent remonter jusqu'au soma et informer la cellule sur ce qu'il se passe à la périphérie ou jouer un rôle trophique (NGF) (communication cellulaire). LE TRANSPORT DES MITOCHONDRIESIl consiste en un transport des mitochondries nouvellement formées dans le soma jusqu'aux terminaisons axonales à une vitesse de 10 à 40 mm par jour. Ce transport consiste en des mouvements saccadés de va-et-vient. Les mitochondries en dégénérescence (corps multilamellaires) remontent vers le corps cellulaire pour y être dégradées. Le mécanisme intime de ce transport reste inconnu. | ||||||||||
|
Iconographie personnelle - Dr. D. Rose | ||||||||||
|
Le neurone est une cellule excitable et sécrétrice qui présente une extrême régionalisation de ses fonctions :
|
Inscrivez-vous au blog
Soyez prévenu par email des prochaines mises à jour
Rejoignez les 77 autres membres